研究小组阐明了自古以来植物形成次生芽的系统
一个合作研究小组已成功鉴定出重要的转录因子GCAM1,该因子可通过创建克隆后代(无性繁殖)来使艾蒿植物无性繁殖。此外,该转录因子被揭示与被子植物中调节次级芽形成的那些转录因子具有相同的起源。这种转录因子是在艾蒿中发现的,这表明它至少是4.3亿年前在陆地植物共同祖先中的一项重要的古代发展。
希望这些发现将导致技术的发展,以提高农业和园艺业中各种植物的种植率。
该小组由神户大学理学研究科的研究员安井幸子(现为京都大学助理教授)和石崎副教授组成,此外还包括京都,信州和近代大学以及马克斯·普朗克植物育种研究所的成员。 (MPIZ)。
分生组织是位于可能发生生长的植物区域(通常在植物尖端)的未分化或未专门化的细胞库。这些分生组织可以使新芽和花朵生长,此外还可以通过创建自身的克隆使许多植物品种无性繁殖。此过程称为营养繁殖。通过营养繁殖形成的克隆后代具有与亲本植物相同的遗传组成。营养繁殖发生在不同地区,具体取决于植物的种类,例如茎(马铃薯,草莓),根(红薯)或什至叶片(kalanchoe)。也可以使用许多技术来诱导它,例如插条和嫁接,并且通常用于在农业和园艺中生产植物。然而,
图2:GCAM1突变体的外观。上图:三周大的地艾植物。左侧是常规野生型艾蒿(用作对照),右侧是突变的艾蒿,其中已去除了GCAM1基因。下图:宝石杯的横截面。红色箭头表示气室,蓝色箭头表示腹部。在突变体中(右侧),没有形成宝石杯。信用:神户大学
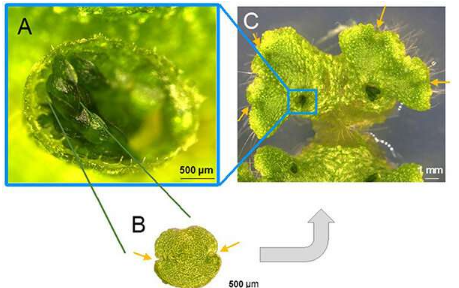
地草的营养繁殖发生在植物体表面的宝石杯内(图1)。一个由一些相同成员组成的研究小组最近阐明了宝石在宝石杯中发育的机制。但是,目前尚不清楚什么机制可以使宝石杯最初形成。
该研究小组试图通过对基因表达的全面分析来阐明哪些基因在宝石杯的形成中起着重要作用。通过这项分析,他们找到了一个转录因子,称为与Gemma杯相关的MYB1(GCAM1)。他们发现,从植物中去除该GCAM1基因会阻止形成宝石杯(GCAM1突变体-图2)。然而,除宝石杯外,该基因的去除并没有影响植物生长的其他方面,导致他们得出结论,编码GCAM1基因的GCAM1蛋白对于触发宝石杯的形成至关重要。
接下来,他们阐明了GCAM1蛋白的功能,旨在控制宝石杯的形成。为此,他们在艾蒿植物体内产生了GCAM1蛋白的过表达。当GCAM1的蛋白质功能过度表达时,植物将无法生长,具有干细胞特性的未分化细胞也会增加(图3)。基于该结果,认为GCAM1负责抑制分生组织区域中的细胞分化,这在正常条件下将允许形成宝石杯和宝石。
GCAM1是R2R3-MYB转录因子基因。已知被子植物中相同的基因亚家族具有腋芽形成功能(拟南芥中的RAX和番茄植物中的盲)。为了研究这种进化关系,使用模型植物拟南芥进行了实验。
先前已经发现,与野生型(对照)植物相比,没有RAXs基因的突变型拟南芥植物几乎没有腋芽。但是,该研究小组发现,将GCAM1基因引入该突变体会导致腋芽的数量增加。利福草的GCAM1基因在拟南芥中显示出相同的功能,表明RAXs基因和GCAM1基因是直系同源基因(图4)。直系同源物是不同物种中的基因,它们是从共同的祖先基因进化而来的,因此保留了相同的功能。
结果表明,地草的GCAM1基因和被子植物中腋芽形成的基因是直系同源物。根据该发现,推测地艾中的胚芽杯形成与被子植物中的腋芽形成具有相同的机理。
控制被子植物腋芽形成的技术可用于直接提高农作物的产量。希望对GCAM1的进一步研究可以为此类技术奠定基础。接下来,有必要阐明参与GCAM1管理的网络,并研究地草和被子植物之间的共性。这不仅有助于了解地艾的营养繁殖,而且还将揭示植物产生新芽的常见机制。